Carbohidratos
Biomoléculas de valor energético y estructural, las más abundantes.Tienen varias funciones, desde energéticas a señales de localización intracelular (clicocálix). Son principalmente, polihidroxialdehidos y polihidroxicetonas. Solo algunos azúcares estan formados por N, P o S. Su fórmula empírica es (CH2O)n. Se dividen en:
Monosacáridos:
Subunidad menor de los carbohidratos. Su nomenclatura tiene relación al número de C que presentan:
3C = triosa
4C = tetrosa
5C = pentosa
Noten que todas terminan en OSA
Los monosacáridos no son hidrolizables y a partir de 7C se vuelven inestables. Tienen un esqueleto carbonado con grupos alcohol o hidroxilo y son portadores del grupo aldehído (aldosas) o cetónico (cetosas). Son solubles en agua, dulces, cristalinos y blancos. También presentan la característica de esteroisómeros (imagen de espejo) y existe solo un azúcar que no lo es, la cetotiosa o dihidrocetona.
La disposición del grupo -OH a la derecha en el C asimétrico determina el isómero D, si está situado a la izquierda es un isómero L. Cuando un monosacárido tiene varios esteroisómeros, todos los que poseen a la derecha el grupo OH del C más alejado del grupo carbonilo son de la serie D, y los que lo poseen a la izquierda son L.
Si dos monosacáridos se diferencian solo en el -OH de un carbono se denominan epímeros. Si son imagenes o reflejos de un espejo entre sí se denominan enantiomeros.
Si un monosacárido tiene más de 3C empieza a tener más de 1C asimétrico. El número de C estereoisómeros es igual a 2 elevado a n, siendo n el número de carbonos del monosacárido. Para una hexosa, el número de estereoisómeros es de 16, de los cuales la mitad, o sea 8C son de forma D y 8 de forma L
La mayoria de los monosacáridos sintetizados por los organismos vivos son D isómeros debido a la especificidad de las enzimas que los sintetizan.
Los monosacáridos son inestables por naturaleza, para estabilizarse, se ciclan, para esto los grupos aldehídos o cetonas reaccionar con un hidroxilo de la misma molécula convirtiéndola en anillo. Las hexosas se ciclan en anillo piranóbico y las pentosas en anillo furanóbico.

Cada azúcar tiene su equilibrio con
respecto al tipo de anillo que se forma, ya sean ![]() o
o ![]() . En el caso de
la Glucosa, la forma
. En el caso de
la Glucosa, la forma ![]() -D-glucosa lo
hace en un 2/3 de ka solución, mientras que la forma
-D-glucosa lo
hace en un 2/3 de ka solución, mientras que la forma ![]() -D-glucosa lo
hace en 1/3. Además, hay una pequeñisima parte en forma lineal.
-D-glucosa lo
hace en 1/3. Además, hay una pequeñisima parte en forma lineal.
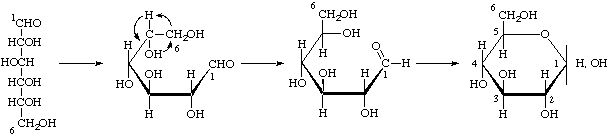
Con las formas cíclicas aparece un nuevo carbono asimétrico o anómero (el que antes tenia el aldehído o cetona).
Los anómeros serán ![]() si el -OH de
este nuevo carbono asimétrico queda hacia abajo y
si el -OH de
este nuevo carbono asimétrico queda hacia abajo y ![]() si lo hace
hacia arriba en la forma cíclica. Los anillos que se forman no
son planos, por lo que pueden adoptar dos formas, la de
"cis" o la "trans"
si lo hace
hacia arriba en la forma cíclica. Los anillos que se forman no
son planos, por lo que pueden adoptar dos formas, la de
"cis" o la "trans"
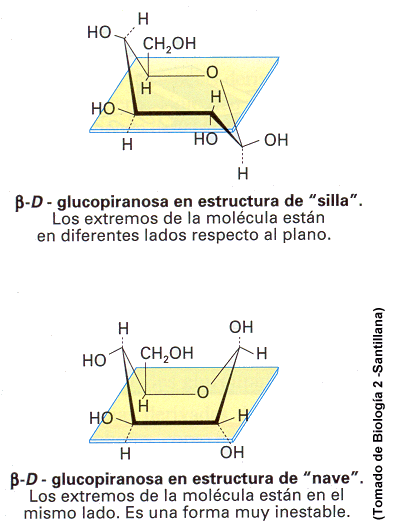
La mayoria de los monosacáridos simples, de 3 a 7 C, son dulces.
Se puede demostrar que la forma cíclica está en equilibrio con la forma lineal de un azúcar en solución acuosa, ya que la forma lineal puede ser oxidada por iones Fe+³ o Cu+² de aldehido a ácido carboxilo. Esta propiedad se denomina podre reductor de los azúcares y la tienen los monosacáridos y oligosacáridos, con entremos hemiacetálicos o hemicetálicos libres.
Derivados de monosacáridos:
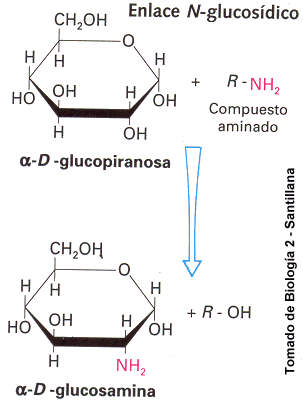
Los principales azúcares derivado son hexosas, que tienen un grupo hidroxilo sustituido por otro químico o un átomo de carbono oxidado a un ácido carboxílico. El primer tipo de derivados de azúcares son los que tienen el grupo hidroxilo del C2 reemplazado por un grupo amino (NH2) Ej: glucosamina, galactosamina.
Existen varios derivados de la glucosamina que tienen el grupo amino del C2 condensado con ácido acético y forman la N-acetilglucosamina, que es constituyente de la pared bacteriana. Otro derivado, también presente en la pared bacteriana, es el ácido murámico en el que el hidroxilo del C3 está condensado con ácido láctico y el ácido N-acetilmurámico, además, el grupo NH2 del C2 está condensado con ácido acético.
Otro tipo de derivado de hexosas son las deoxiazúcares, como la fructosa y ramosa, que son derivados de la galactosa y manosa, respectivamente. En estas deoxiazúcares el hidroxilo del C6 esta reducido a H.
CH2OH--->CH3
Estas deoxiazúcares forman parte de las glicoproteínas y glicolípidos.
Existen tambien los azúcares ácidos en los que el grupo hidroxilo del C1 está oxidado a ácido carboxílico y forman ácidos aldónicos y los otros son los que tienen el grupo aldehido del C6 oxidado a ácido carboxílico para formar ácido urñanicos. Ambos tipos de ácidos forman ésteres intramoleculares que son estables, denominados lactonas. Ej: Existe un ázucar ácido derivado del ácido de la N-acetilmanosamina llamado ácido N-acetilneurominico o ácido siálico y se encuentra presente en muchas glicoproteínas y glicolípidos
Además, estan los azúcar fosfato, en los que las hexosas están condensadas con ácido fosfórico en el C6. Este tipo de derivados de hexosas son comunmente intermediarios metabólicos en los que el grupo fosfato activa el azúcar para que sea más reactivo y puede reaccionar con otras moléculas. Además, estos derivados fosfatados de azúcares no traspasan las membranas a menos que haya un transportados para ellas.
Oligosacáridos
Cadenas de monosacáridos de 2 hasta 100 azúcares. Su forma más abundante es como disacárido.
Polisacáridos
Cadenas largas de monosacáridos, de varios cientos o miles y que pueden ser ramificados o lineales. Los polisacáridos, a diferencia de las proteínas. no tienen un peso molecular definido, ya que no son sintetizadas a partir de un molde (RNAm) como las proteínas.
Las que determinan el peso de un polisacárido son las enzimas responsables de todos los pasos de la síntesis que actúan secuencialmente. Existe una enzima para cada tipo de unión de cada monosacárido diferente.
Los mecanismos que determinan el peso máximo de un polisacarido son desconocidos.
Tanto el almidón, que pertenece a las células vegetales, como el glicógeno, de las célular animales, son pilisacáridos de almacenamiento que se acumulan formando gránulos. Estos polisacáridos están altamente hidratados ya que tienen cientos o miles de grupos OH expuestos al medio acuoso. Ambos son polímeros de glucosa en distintas estructuras.
El almidón se acumula principalmente en tubérculos y semillas de plantas, está compuesto por 2 tipos de polímeros de glucosa:
El glicógeno es un polímero ramificado de glucosa como la amilopectina del almidón, salvo que las ramificaciones ocurren cada 8-12 residuos en la cadena lineal por lo que es un polímero más compacto que el almidón.
En las células animales, el glicógeno se encuentra en forma de gránulos en ciertas célular como las hepáticas y musculares. Cada molécula de glicógeno tiene un peso de varios millones y tiene asociado a la molécula las enzimas de síntesis y degradación del polímero.
Tanto para degradar la amilopectina como el glicógeno, muchas enzimas especificas se unen a los extremos no reductores y actúan al mismo tiempo.
La glucosa es almacenada al interior de la célula vegetal y animal como un polímero de alto peso molecular, y no como glucosa libre, ya que de esta forma contribuye poco a generar diferencia osmolar entre el interior de la célula y el medio externo (la osmolaridad está dada por el número de moléculas en solución y su diferencia con el medio externo).
Celulosa y quitina.
Tanto la celulosa como la quitina son homo polisacáridos estructurales. La celulosa es un polímero de D-glucosa unidas por enlace glicosídico B1-4 y la quitina un polímero de N-acetilglucosamina con el mismo enlace.
el enlace glicosídico b1-4 genera polímeros linealesmás rígidos que los de la amilosa que son A1-4 y permite que varios polímeros lineales interactúen entre ellos formando fibras que son más resistentes.
La celulosa forma la parte leñosa y resistente de muchos vegetales, la quitina compone la caparazón de muchos artrópodos.
Los animales pueden degradar la amilosa, amilopectina y glicógeno, ya que tienen enzima A-amilasa, que hidroliza el enlace A1-4, pero no pueden degradar celulosa o quitina porque no tienen enzimas para el enlace B1-4. Solo los rumiantes (vacas, ovejas, jirafas, etc), que tienen un estómago extra llamado Rumen, en el que viven bacterias y protozoos, pueden degradar la celulosa por las celulasa secretada por estos.
Los glicosaminoglicanes y protroglicanes de la matriz extracelular.
Los glicosaminoglicanes y protroglicanes son heteropolisacáridos que conforman la matriz extracelulas que existe entre las células de los tejidos animales y que le atorga la consistencia de gel.
Los glicosaminoglicanes son polímeros lineales compuestos por la repetición de un disacárido. el disacárido es N- acetilglucosamina o N-acetilgalactosamina que está unida a un ácido urónico, muchas veces, ácido glucurónico. Los grupos OH de la N-acetilglucosamina o galactosamina están muchas veces esteficados con un grupo fosfato y sumado a los grupos carboxilo del ácido urónico le dan carga negativa al glicosaminoglicano (=forma extendida en solución)
La forma extendida de los glicosaminoglicanes le da viscosidad a la matriz extracelular.
Existen tres tipos principales de glicosaminoglicanes:
Los proteoglicanos están compuestos por una molécula central larga de ácido hialorónico a la que se unen de forma no covalente, pequeñas proteínas (20.000) y a las que se unen de forma covalente otros glicosaminoglicanos como el condratán sulfato, el queratín sulfato, hiparán sulfato y dermatán sulfato.
Los glicosaminoglicanes antes mencionados están unidos covalentemente a una proteína a través de residuos de serina de las proteínas y tienen en genral forma de árbol cuando el conjunto de ellos se acopla a la forma central del ácido hialurónico.
Los proteoglicanos pueden llegar a ser moléculas inmensas que se entremezclan con las fibras de colágeno y elastina para formar la matriz extracelular.Se unen a ciertas proteínas de membrana, y estas se unen a su vez a proteínas integrales de membrana llamadas integrinas. De esta forma se anclan los proteoglicanos de la matriz extracelular a la célula.
Las glicoproteínas
La mayoría de las proteínas que son secretadas por las células eucariótica son glicoproteínas.
En las glicoproteínas, la proporción de azúcares, comparado con la proteína, es de 1 a 70 (menos que en los protroglicanos)
La cadena de azúcares unidos a una proteína le da una gran carga negativa en la razón de unión y la cadena de azúcares toma una forma mas bien lineal y extendida que protege a la proteína del ataque de enzimas proteolítica.
Los oligosacáridos unidos a proteínas son generalmente heterooligosacáridos, en los que se combinan diferentes monosacáridos con diferentes tipos de uniones (1-3, 1-4. 1-2, 1-6, 2-3) y en los que los monosacáridos están en conformación A o 2.
Puede haber varias cadenas de oligosacáridos unidos a una misma proteína. Las cadenas pueden ser cortas o largas, y lineales o ramificadas.
Los oligosacáridos se unen a la proteína a través de un enlace O-glicosídico cuando el azúcar terminal se une al OH de una serina o treonina, o a un enlace N-glicosídico cuando el azúcar se una a una asparagina.
La enorme cantidad de azúcares que existen, le dan una variabilidad muy grande a las glicoproteínas. Estas cadenas de oligosacáridos deben también ser reconocidas por las enzimas y receptores que interactúan con la glicoproteína. Al parecer, estas complejas cadenas de oligosacáridos codifican para información que determina la estructura terciaria de la proteína, la ubicación de ella dentro d la´célula y la manera en que será reconocida por una enzima o receptor.
Enlace glucosídico
Existen dos tipos de enlaces glucosídicos, el O y el N
Enlace O-glucosídico:
Los monosacáridos de unen a través de este enlace, que se realiza entre el grupo hidroxilo del monosacárido 1 y el carbono anomérico del monosacárido 2, deshidratandose ambos, al formar una molécula de agua.
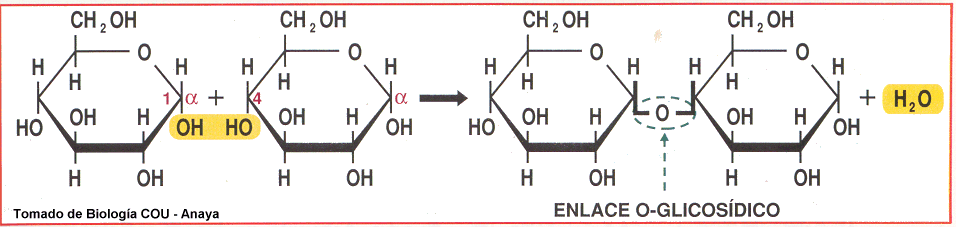
Las uniones glicosídicas pueden ser hidrolizadas por ácidos débiles en caliente. El C anomérico de un azúcar puede reaccionar con un átoma de N para formar un enlace N-glicóndico como en el caso de los nucleótidos.
Enlace N-Glucosídico
Realizado entre un grupo OH y un compuesto aminado, para formar aminoazúcares