The
Blue Question
Richard
Cryberg
February
14, 2012
Introduction:
Blue
in pigeons is defined as a part of what we refer to as wild type.
Therefore, it is of some interest to consider both the
history of wild type, and the occasional deviations and or
misinterpretations that arose as our understanding of
genetic science developed. It is also of interest to
understand what
physical phenomena lead to what we call a blue color and examine the
history of
how this thinking evolved. Just so we are all on
the same page, I show in Picture 1 below, two blue
secondary flight feathers.
Picture
1: Left
feather appears blue barless and right feather is from a blue bar.
 Note
the grainy transition between blue and black areas on
the barred feather. This grainy
transition is why Hollander named this type of black coarse
spread.
The coarse refers to the naked eye nature of
this transition. Other apt words he
might have used would be granular transition spread as the transition
has a
naked eye granular look. I will address
later why the left feather is labeled apparent barless. I
will show that pigment clumping does not exist and that this
so called clumping has nothing to do with production of what we call a
blue
feather.
Note
the grainy transition between blue and black areas on
the barred feather. This grainy
transition is why Hollander named this type of black coarse
spread.
The coarse refers to the naked eye nature of
this transition. Other apt words he
might have used would be granular transition spread as the transition
has a
naked eye granular look. I will address
later why the left feather is labeled apparent barless. I
will show that pigment clumping does not exist and that this
so called clumping has nothing to do with production of what we call a
blue
feather.
Discussion :
Wild
type is the sole basis for all of biology. It was well
established long before Darwin or Mendel were even born, much
less before the field of genetics began to be understood in even a
minor way. It is still today the sole basis for making
sense and order in biology. In fact, the
rudiments of the wild type idea probably trace back for 1000 years or
more. It is pretty hard to pin down an
exact date when it was first recognized, nor who the first person was
who
recognized it. If you do not understand
wild type you do not understand anything about biology that has
happened for at
least the last 300 years.
The
first person to formalize wild type was Carl Linneaus in
the middle 1700s. Linneaus recognized
that plant and animal species differed from each other in measurable
physical
characteristics. A given species
differed in physical characteristics within that species far less than
those
same physical characteristics differed from other species.
Based on this observation he developed a
classification scheme that is still in use today.
Today’s version follows an order where each
step down represents a group that is more and more closely
related.
The steps are Life, Domain, Kingdom, Phylum,
Class, Order, Family, Genus, Species.
This is something taught in high school biology. At the end,
species, you are talking about
one single species. One level up, genus,
you are talking about a number of species which are closely related
based on
phenotypic similarity.
It
is not always an easy decision if two populations that
are very closely related are one species or two species. In
pigeons, for instance, some have argued at
times that wild populations that were checked were a different species
than
wild populations that are barred. In
fact variation in wild population from one location to another location
is
perfectly normal and to be expected. So
biology has people who are called lumpers and others who are called
splitters. A lumper would consider
barred and checked populations of pigeons as one species. A
splitter would call them two species. The lumper and splitter
disagreement is
unending, even today, in some cases. In
general though, this disagreement is eventually resolved on one side or
the
other. Often today. things like DNA
sequence data are brought to the table as part of the argument; as well
as
observations on interbreeding and gradation in phenotype in the
geographic
areas between the two different population centers. Today we
also realize, that at times
phenotypes can be so close to identical, that two species can be
difficult to
tell apart simply by physical examination.
So DNA evidence is becoming more important all the time.
This
idea that members of a species share more phenotypic
traits with each other than they share with any other species was very
well
established by Darwin’s time in the middle 1800s; and formed
a considerable
amount of the background information he used to formulate his ideas
about how
one species evolved into another species.
Darwin’s thought was if a population of one species was
geographically
isolated from other members of the same species that isolated
population would
live under unique conditions and that with time it would evolve traits
that
allowed it to better survive in its local environment. What
Darwin recognized was wild type was not
forever stable but could change as a result of what he called survival
of the
fittest. Like most new ideas in science
this idea was not warmly embraced by some people. A great
deal of his thinking was in fact
based on domestic pigeons. But, a great
deal was also based on his observations of finches on the Galapagos
Islands. Each island had finches which
were different in some specific ways from finches on the other
islands.
Often beak sizes and shapes differed
dramatically. We know today these beak
differences are not even due to differing genes. They are
caused by changes in switches that
turn genes on and off. Darwin recognized
that these finches were all very closely related and probably all
evolved from
one single finch species. He was
correct.
In
the case of domestic pigeons, man interfered with nature
and selected for physical traits man found desirable. The
result was different breeds that had
greatly different phenotypes. Yet he
knew these different breeds would mate and produce young which were
perfectly
vigorous and fertile. And further these
crosses generally looked much more like the well recognized wild type
for
pigeons than either parent breed. Darwin
had no clue at all how this was possible.
He came to these conclusions many years before Mendel’s work
on peas
became available. About all that was
well understood at Darwin’s time in domestic animals was if
you selected for
some given trait you could over time develop that trait and exaggerate
the
difference from wild type. In fact, every
domestic animal and plant was recognized as having been so selected by
man to
the point that in some cases it was not obvious what the original
ancestor wild
species had been.
Gregor
Mendel published a paper in
1865
about peas concerning what
we today call inheritance; and it became instantly ignored.
In the next 35 years, only three scientists
even referenced his work. The thinking
at that time was that inheritance was a blending operation of some sort
in
spite of Darwin’s observations on pigeons which clearly
refuted any such
idea. It seems the idea that there could
be units that were passed from parent to offspring that survived intact
without
dilution was more than biology at that time could accept. The
net result was Mendel’s work lay dead
until 1900.
Hugo
de Vries and Carl Correns rediscovered Mendel’s idea of
unit inheritance in 1900 and rediscovered Mendel’s earlier
papers and gave him
proper credit. Not uncommonly, in science,
an idea is originated that is simply before its time and lays dormant
for many
years. That is exactly what happened to
Mendel. Suddenly in 1900 the study of
what we now call genetics exploded. As
is typical in any explosion this new science instantly became
disorganized and
fragmented and filled with many ideas that today seem insane.
The early students of genetics largely lost
the idea of wild type or at least made a pigs ear of it. Much
of the reason this happened is
understandable today and probably was inevitable.
Understanding of statistics was rudimentary
at that time except when applied to games of chance and nonexistent in
the
biological sciences. There was no clue
at all what these units of inheritance were or where they
resided.
Biochemistry knowledge consisted mainly of
how to make bread, beer, wine and booze and not much more.
And, even with bread, beer, wine and booze
the physical process was understood but not much that was happening on
a
molecular level.
The
net result was in early papers on genetics some things
were proposed that today we view as amazing and can not understand how
such a
thing could possibly have ever been suggested.
It took many years of experimental breeding programs carefully
gathering
data before it became obvious that things like the color of a pigeon
involved a
whole host of wild type genes and alteration of any single wild type
gene could
lead to a new phenotype. Early on the
thinking was more along the line that color, or any other phenotypic
trait in
pigeons was probably the result of only one or two genes. The
lack of understanding statistics also
lead people to make some mating, raise three young and draw conclusions
as to
some trait being dominant or recessive.
It is easy to understand such errors when you realize that the general
wisdom was most of the time only one gene was involved in any kind of
trait. For example, with the thought
process at the time all crests on pigeons could well be considered due
to
different variations of one single gene that when mutated gave crest
phenotypes. Or all the different forms
of feathered feet and legs could be due to variations in one single
gene that
when mutated in different ways gave various feather patterns and sizes.
The
first paper on inheritance in pigeons of any
significance was published by Leon Cole in 19141.
In that paper he stated “Red may be called
the fundamental color in pigeons as is probably the case in fowls and
most
mammals. It appears to be potentially
always present, and if we let the factor for its production be
represented by
the letter R, we may say that apparently the factor R is never lacking
from the
gametic formula of pigeons.” The red to
which Cole refers is what we today call recessive red. In two
prominent, short sentences Cole discards
the whole concept of wild type that was the basis of all of biology for
the
prior 200 plus odd years! It was another
decade or decade and a half before the science of genetics had made the
progress to understand that there were three total disasters in
Cole’s two
short sentences. He not only discarded
wild type but far worse he gave a unitary name and symbol to his newly
defined
wild type. It is not clear at all why he
happened to assign R rather than r to red.
It is clear he understood dominants and recessives. He
assigned S to spread and B for blue for
example. His gametic formula for a red
was RRbbSS so he recognized that good reds at times have spread as one
component. His formula for blue was
RRBBss and black was RRBBSS. It is
pretty hard to understand how in the world errors of this nature could
possibly
have ever happened unless you study history.
If you study history you will learn that such errors were inevitable
and
are simply a pretty normal, although discouraging, part of how any
science
develops over time. When you are
operating with very fragmented knowledge and only have tiny bits and
pieces of
actual data to study and think about and when you have not allowed
enough time
for other fields of science to weigh in and cast a vote, errors happen.
This was
true in the case of genetics in 1914 as genetics was only a 15 year old
branch
of biology at that time. The net result
is a pretty normal high school student today can easily have a much
more
sophisticated understanding of genetics than the best living scientist
had in
1914. We simply have to remember that in
1915 no one had any clue what a gene was or even where in the cell
genes were
located. No one had discovered DNA. A perfectly
reasonable person in 1915 could
have thought a gene was simply some bit of protein or carbohydrate that
floated
around in the cells cytoplasm. We Also
must remember that in 1914 there was no reason to realize, as we
clearly
realize today, that for every mutant gene there exists a wild type
gene.
And, at least for recessive mutant genes that
wild type gene does things that are absolutely critical in making a
wild type
phenotype. It was not at all understood
back then that if you removed the wild type form of the recessive red
gene you
would not get a blue pigeon. This is so
clearly understood today that a high school kid can understand the
reasons.
But,
in
1914 it simply was not
understood at all. Even today, almost no pigeon
persons understands what would happen if the wild type
form of
the recessive red gene was removed. Thus it is reasonable to
give Cole a lot of
forgiveness.
You
could make exactly the same type of disparaging
observations
about Newton failing to recognize the theory of relativity
for
example. After all, he had every single
tool available needed to recognize relativity as inevitable.
In fact, he knew every physical principal and
the needed math. He knew the math far
better than Einstein knew it; as Newton invented the needed math and
Einstein
was a miserable math student and learned little in school about the
topic. Newton was in a vastly better position to recognize
relativity back when he did his work, than Cole was in, to understand
pigeon
genetics in 1914. The wonderful part of
science is it has a built in self correcting process that takes care of
such
errors with time. That is why science
demands disclosure of all relevant data when a paper is
published.
With data, it is possible for another worker
to go back and not only replicate experiments but also do added tests
to
confirm or deny claimed results as well as explore new
possibilities.
Thus the statement, no data, no science.
Unfortunately,
experience has taught me that most of the
pigeon hobby has a current understanding level such that they still
largely
fail to understand wild type. There
likely is no cure as science had corrected Cole’s wild type
errors within a
decade, yet many people in the hobby still talk about a blue
gene.
They totally ignore the fact that a blue gene
is forbidden by the naming rules because there are so many genes that
equally
deserve the name blue gene.
The
corrections to Cole’s errors came more from fruit fly
studies than any other single place.
Fruit fly studies showed that any given phenotypic trait is frequently
impacted by mutants at multiple different locations. In some
cases, mutants at two clearly totally
different locations could lead to exactly the same phenotype or at
least so
close it can be hard to tell the two mutants apart simply by looking at
phenotype. This forced the genetics
community to regroup and go back to the old standards that biology had
accepted
for centuries, namely wild type. And
part of going back was to forbid the practice of giving wild type genes
for a
phenotype due to the vast confusion that resulted. This
happened in the 1930s. As we have come to understand the
biochemistry involved in heredity and life; it has simply become more
obvious
that naming or symbolizing a wild type gene based on phenotype is a
fool’s
errand. Today’s rule is simple and
unambiguous. Wild type is named for the
first discovered mutant at that locus.
For example, wild type at brown is a proper name for a particular wild
type gene in pigeons. Other examples
would be wild type at dilute, wild type at recessive red,
etc. The exception is if the exact biochemical
function of that wild type gene becomes known and the gene’s
DNA has been
sequenced. If both criteria are
satisfied it is perfectly acceptable to name it for its biochemical
function. For example the wild type at
albino gene is also properly named the tyrosinase gene as tyrosinase is
the
enzyme that wild type gene produces. It
is symbolized Try. Both biochemical
function and DNA sequence are absolute requirements for such a
name. Absent either no name is permitted.
Cole
made other errors in his 1914 paper. A major error he made
concerned blue. Cole never looked at pigeon feathers under
the microscope. Rather, he states
clearly that he is accepting the unpublished findings of Lloyd-Jones as
his
source of information. Lloyd-Jones
published his paper2
in 1915.
As Lloyd-Jones was in Iowa and Cole in Rhode Island in those days
probably all communication between them was by letter.
Further, Lloyd-Jones did not have
photographic equipment available so he had to hand draw things he
viewed in the
microscope. Between slow communications
and hand drawings there was much opportunity for errors to creep into
the
understanding of what pigment granules look like and how they are
distributed
within the feather. Cole was the first
to state that pigment granules are clumped in blue feathers.
He states that what we call blue “is in
reality not a blue but a neutral shade of gray.
It corresponds to Ridgeway’s ‘gull
gray.’
The optical effect is due to a different
arrangement of the pigment in the barbules from that which obtains in
blacks. In blues the pigment is
aggregated into clumps, while in blacks it is spread uniformly
throughout the
barbules.”
I
am sure if Cole had ever actually looked at a blue feather
under a microscope even at 100X magnifications he would never have said
something so foolish. This is not even
approximately true of a blue feather.
Nor is it what Lloyd-Jones reported in his paper a year later entirely
accurate. Yet, from 1914 to today the
idea has persisted that somehow pigment clumping has something to do
with
production of the blue in a pigeon feather.
It seems I must be the first person since Lloyd-Jones to actually look
at blue pigeon feathers under a microscope at a variety of
magnifications! Clumping does not exist. It is an
invention by people who do not have
any idea what the distributions of pigment actually looks like because
they
have not looked at feathers themselves with a microscope and because
they have
not read Lloyd-Jones 1915 paper carefully nor my paper published on
Huntley’s
web site3.
Unfortunately
Lloyd-Jones picked a sample preparation
technique that lead to considerable loss of detailed data at lower
magnifications. His sample preparation
method involved cutting feathers apart with scissors, embedding the
jumbled
parts in wax then cutting thin sections with a microtome. I
am skipping a number of staining and
clearing steps he also took. The problem
with this sample preparation method is he could not always tell exactly
what
part of the feather he was looking at and could not scan the scope
across a
feather with a traveling stage and get an overall view. He
also concentrated his main effort of the
smaller parts of the feather such as the barbules and barbicels and
largely
ignored the larger parts such as rachis and barbs. In his
paper he states concerning a blue
feather that “In the barb the pigment is restricted entirely
to the apex. The lateral sheets of cortex are altogether
without pigment and the medullary cells, richly supplied with pigment
in black
or duns, are entirely without pigment.”
I can not understand how he could possibly have drawn this
conclusion. In picture 2 I show a single
barb with its attached barbules and barbicels.
The barb is the large black central shaft in this picture that goes
clear from the bottom to the top of the view.
Picture
2. Blue at 100X
magnification
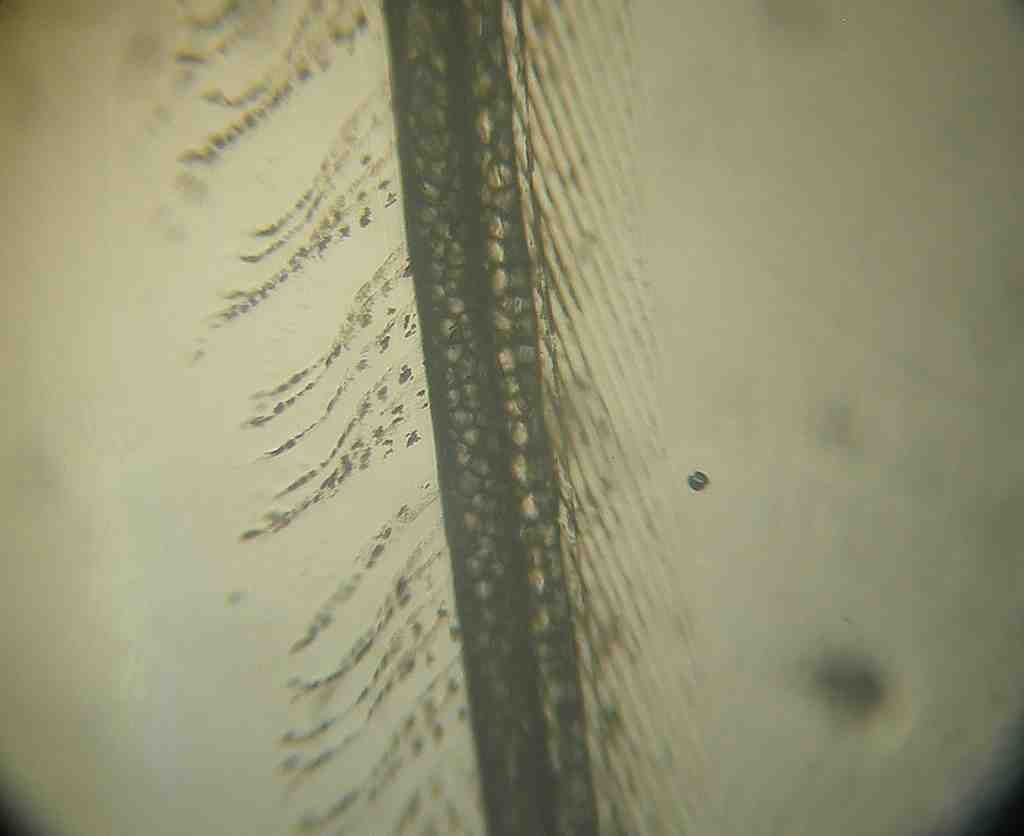
The
first thing to note is practically all the pigment is
located in the barb shaft itself in contrast to what Lloyd-Jones
reported. A recessive white barb shaft shows no hint of
color at all so the color seen in a barb from a blue feather is due to
a lot of
pigment and not just a lot of material preventing light
transmission. And rather than being restricted entirely to
the apex this pigment is distributed along the whole barb
shaft. I have absolutely no explanation for why
Lloyd-Jones said he saw what he reported other than perhaps his sample
preparation technique made such a mess of the feather parts he did not
always
understand what particular part he was looking at. At any
rate, it is obvious from picture 2
that most of the total pigment is in the barb shaft. My crude
estimate is that over 95% of the
total pigment present in a blue feather is in the uniformly pigmented
barb
shaft. This estimate is much more obvious
when looking at the sample under the microscope than it is looking at a
picture. Under any circumstances the
amount in the barbules is a minor part of the total pigment
present. Lloyd-Jones speculates that the color of a
blue feather is somehow influenced by the non-uniform distribution of
pigment
granules in the barbules. He does use
the word clumped a few times to describe the distribution of these
granules in
the barbules. He is also careful to give
both other word descriptions of the arrangement of these granules and
has some
excellent hand drawings of the distribution.
These granules in the barbules are not clumped. The fact he
used the word clumped is
unfortunate but likely simply the result of fishing for alternate words
to use
to avoid over use of other words.
Pigment granules in these islands or groups are clearly resolvable as
single granules at high magnifications with no contact with their
neighbors. In general any two granules
are separated by one or more granule diameters.
This
kind of distribution does not fit any normal definition of
clumped. Groupings of granules, Yes. Islands of
granules,
Yes. Clumps of
granules, No. So
forget that anyone ever used
the word clumped to describe this pigment distribution as the word
clumped
simply does not fit reality.
Distributing these same
granules found in the barbules uniformly in the
feather is going to have no significant impact on the color of the
feather. In fact, if that were the only
pigment in the feather the color you saw would be near white regardless
if the
pigment were in islands or uniformly distributed.
It
is obvious that when Cole wrote his paper he simply relied on
what Lloyd-Jones was telling him
could be seen with a microscope. So Cole
decided that black was due to some genetics that resulted in
distributing the
exact same pigment granules spread uniformly though the feather
parts.
Lloyd-Jones had also looked at such birds and
reported the feathers were colored in all parts. That was the
origin of the word spread. It is curious that Cole seems to
have ignored
entirely Lloyd-Jones observation that pigment granules varied in size
in some
cases and granules in blue feathers were gigantic compared to granules
in a
spread black pigeon. In fact it is clear
that by the time Lloyd-Jones published his paper a year after
Cole’s paper he
was not at all comfortable with Cole’s
interpretation.
In particular Lloyd-Jones did not like Cole’s
use of the word spread at all. He wrote
“It seems wise to the writer to retain the symbol S in the
genetic formula for
pigeons, but with this modification, that we consider it a factor for
‘stopping’ rather than for
‘spreading.’”
It is clear that Lloyd-Jones realizes that a great deal more than
simply
pigment distribution within the feather is involved in turning blue
into black,
including the fact that blacks simply having a great deal more total
pigment
and a different size of pigment granule.
The
conflicts between these two workers is rather surprising
as they were obviously communicating their findings to each
other.
I can only surmise that Cole did not give
Lloyd-Jones the courtesy of reviewing his paper before
publication.
Had this happened a lot of future confusion
that has continued to this day might have been avoided.
Namely that clumping of pigment granules had
something to do with production of what we call the blue color of
pigeon
feathers. Still, they would not have
gotten the story right as they missed all the pigment in the barb shaft
itself
and concentrated their effort looking at the wrong part of the feather.
More
recently we have the quantitative data on amount of
pigment present in blue vs. black. Sell,
et all4
report that the blue part of a feather in a spread blue has
nearly ten times as much total pigment as a blue pigeon. It
seems reasonable that ten times as much
pigment would make the feather much blacker in color. In
fact, this should have a great deal larger
impact on color than spreading granules from groups where they are not
even
close to touching, from an optical light standpoint, to some more
uniform
distribution.
At
the start of this paper I showed picture 1 as an example
of two blue feathers. These are
secondary flight feathers and one appears to be barred and the other
barless. The barless feather is the result
of an interesting accident. For another
microscopy project I needed to look at the barbules and barbicels of
some
feathers. The problem was some of the
feathers had so little pigment I simply could not see some things and
getting
photos was impossible. So, I decided to
dye those feathers. But before using up
feathers of interest to the project I
experimented with dying recessive white feathers. The dye I
used was obtained from a local shop
that supplies all kinds of hair treatment products for women.
If you want pink or green hair they had the
products. I picked a Proctor and Gamble
product, Clairol, Beautiful Collection, Jet Black, shade
B22D.
This is a semi permanent dye and is applied
to damp hair simply by spreading it on the hair, waiting 20 minutes for
human
hair, rinsing well and drying. To dye a
feather I wet the feather out using Palm Olive dish washing detergent
and
rinsed the detergent completely from the feather. I damp
dried the feather with absorbent
tissue paper, spread the dye on both sides of the damp feather with a
tooth
pick and waited various lengths of time up to three hours. I
then rinsed any free dye from the feather
under running water and air dried the feather.
In no case did the feather end up any place close to black.
The deepest color was about equal to a fairly
dark dirty blue. In the case of the
feather shown in picture 1 the dye exposure time was 10
minutes.
In picture 3 I show a single barb and
attached barbules from this dyed feather.
Picture
3. Barb and attached barbules of
a black dyed recessive white feather
 Again
the barb is the large central shaft that runs from the
bottom to the top of the view. As can
clearly be seen nearly all the dye taken up by the feather is
concentrated in
the barb itself and very little is deposited in the barbules.
Most of the dye that is deposited in the
barbules is all towards the end 1/3 of the barbule on only one side of
the
barb. The barbules on the other side of
the barb are for practical purposes nearly dye free. It is
plainly obvious that the blue color we
see in both a natural blue feather and a white feather dyed with black
dye is
the result of both pigment and dye being mainly found in the barb shaft
itself
where pigment and dye are both uniformly distributed. There
is no clumping in the barb. The barbules are for practical
purposes near
white.
Again
the barb is the large central shaft that runs from the
bottom to the top of the view. As can
clearly be seen nearly all the dye taken up by the feather is
concentrated in
the barb itself and very little is deposited in the barbules.
Most of the dye that is deposited in the
barbules is all towards the end 1/3 of the barbule on only one side of
the
barb. The barbules on the other side of
the barb are for practical purposes nearly dye free. It is
plainly obvious that the blue color we
see in both a natural blue feather and a white feather dyed with black
dye is
the result of both pigment and dye being mainly found in the barb shaft
itself
where pigment and dye are both uniformly distributed. There
is no clumping in the barb. The barbules are for practical
purposes near
white.
Thus
the last 95 years of
nonsense about blue having something to do with the non uniform
distribution of
pigment in the barbules should be put to its final rest as a totally
failed and
disproved idea.
1. Leon J. Cole, Studies on Inheritance in Pigeons: I.
Hereditary Relations of the Principal Colors. Bulletin 158,
Agricultural
Experiment Station of the Rhode Island State College, pages 311-385,
1914
2.
Orren Lloyd-Jones, Studies on Inheritance in Pigeons: II.
A Microscopical and Chemical Study on the Feather Pigments, The Journal
of
Experimental Biology, 18,
453-509, 1915
3.
Richard Cryberg, Microscopic Examination of the Pigment
Found on Wild Type C.
livia, https://www.angelfire.com/ga/huntleyloft/Pigment.html
4.E.
Haase, S. Ito, A. Sell, and K Wakamatsu, Melanin
Concentrations in Feathers from Wild and Domestic Pigeons, J. of
Heredity, 83,
64-67, 1992
~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.~.
Email the author, Richard Cryberg:
Back To Start Page
Copyright
2011 by Richard Cryberg
Permission is granted to download or copy for non-commercial individual
use only.
The author retains all other rights under copyright.
Ronald
R. Huntley
Web Page Designer
Duncan SC, 29334
phone: (864) 249-0276