The way in which the components of associative learning - classical conditioning and operant conditioning - contribute to learning in the fruit fly Drosophila melanogaster, has been extensively studied by Brembs (2000), Brembs and Heisenberg (2001) and Heisenberg, Wolf and Brembs (2001), in experiments using a flight simulator.
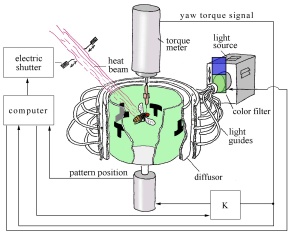
Fruit flies display many kinds of behavioural flexibility in their patterns of bodily movement, or "ethogram". In the experiments, the range of the fruit flies' bodily movements was drastically restricted in a highly unnatural fashion. Each fly, after being immobilized by cold-anaesthesia and glued by the head and thorax to a triangular copper hook, was left overnight with some food (sucrose) in a small moist chamber. The next day, it was attached to a torque meter via a clamp and performed tethered flight in the centre of a cylindrical panorama. The torque meter measured the fly's tendency to turn around its vertical body axis - i.e. its yaw torque. The fly was held horizontally, as if cruising at high speed. The head was glued to the thorax. The fly was unable to turn or shift its gaze or its orientation, but was free to bend its abdomen, extend its proboscis, beat its wings and move its legs. The torque meter to which the fly was attached measured the difference between the fly's left and right wing movements, and interpreted this as an "intended" side-to-side turn.
Most of the experiments were designed to monitor the flexibility of a single behavioural variable in the fly: yawing (movement from side to side). Fruit flies, when allowed to fly freely, do not fly at just one angle, but spontaneously vary their angle of flying over the entire yaw torque range. Fluctuations in yaw torque may be slow or rapid (jerky body movements, known as "spikes"). Torque spikes, generated when the fly reduces its wing beat amplitude on the side to which it "intends" to turn, enable the fly to make sudden turns to the left or right. In the experimental set-up, when the fly "attempted" to make a side-turn, it did so by generating torque spikes.
The flight simulator used in the experiments was a computer-controlled feedback system in which a fly was allowed to control, by its yaw torque, the rotations of a cylindrical panorama surrounding it. In some experiments, the cylindrical arena was used to present a visual stimulus - either a colour or a pattern (a conditioned stimulus). The fly's tendency to perform left or right turns (yaw torque) during tethered flight was fed into the computer. The computer controlled the background visual display of colors and/or patterns perceived by the fly. Thus although the fly could not actually turn left or right, the rotations of the panorama could be used to provide the fly with artificial sensory feedback from its "intended" turns, which mimicked the feedback it would receive in free flight from turning. It also regulated a heat beam which could be focused onto the fly. The heat beam served as an unconditioned stimulus - in this case, a "negative reinforcer" or "punishment". Thirty seconds of continuous exposure to a heat beam would incinerate the fly. A torque meter measured the fly's angular momentum around its vertical body axis - its yaw torque.
The description of the heat beam used in the experiments as a "punishment" raises the question of whether a fly's ability to be conditioned is grounded in its capacity to experience mental states (e.g. pain and fear). Does the heat hurt the fly? And does the fly fear an aversive stimulus? These questions will be further discussed in chapter 3. For the time being, it should be noted that avoidance behaviour alone cannot serve as an indicator of pain or fear: bacteria, whose behaviour is readily explicable using a goal-centred intentional stance, also avoid aversive stimuli.
Brembs (2000) tested fruit flies in four distinct "learning modes", where they were subjected to different combinations of classical and operant conditioning. Two modes were mono-dimensional tasks, where the fly had to form just one association. In operant conditioning, the association was between its behaviour (B) and a "punishing" unconditioned stimulus (US); in classical conditioning, between a visual pattern on the cylindrical panorama (CS) and the US. The other two modes were composite or multi-dimensional tasks, as the fly had the opportunity to form two associations: either B -> US and CS -> US (switch mode) or B -> CS and CS -> US (flight simulator mode).
Brembs (2000) measured the fly's ability to learn from pure operant conditioning by subjecting the fly to heat when it turned too far to the left or right. In this "yaw torque" mode of learning, the fly learned to avoid heat by restricting its yaw torque range, for example, to the left domain if straying into the right domain was punished. The fly thus formed an association between its behaviour (B) of initiating a right turn, and an unconditioned stimulus (US): heat. However, it had to learn this "blind" - i.e. without the help of any external guidance indicating whether it was flying inside or outside the permitted range. Heisenberg, Wolf and Brembs (2001) comment:
This is a remarkable feat, as ... in no natural situation would continuous turning to one side bring the fly permanently out of a heated zone (2001, p. 1).
In Brembs' classical conditioning experiments, a certain visual pattern (CS) on the cylindrical arena was followed by a punishment (US): the fly was subjected to heat. In this set-up, there was nothing the fly could do to avoid the punishment, but subsequent testing showed that it had learned to associate the pattern (CS) with the punishment (US).
In flight simulator mode (fs-mode), an attempt was made to realistically simulate the changes in the fly's visual field that the fly's turns would cause in free flight, except that the flight simulation was maintained at a constant speed at an undefined altitude. The cylindrical arena was decorated with four black T-shaped patterns of alternating orientation - two upright and two inverted - evenly spaced on the wall so that each quadrant had one T-shape. The arena's angular speed was made proportional to the fly's yaw torque, but the direction of rotation was opposite to that of the fly's turning. By adjusting its yaw torque, the fly was able to stabilise the arena and control the direction of its simulated flight (i.e. fly "straight" relative to the patterns on the wall). The fly was punished by heat (the US) when it flew into a forbidden quadrant, marked with an upright T, and it learned that quadrants associated with an inverted T were safe. In this learning mode, the fly was unable to form a direct association between its behaviour (B) and the heat (US), because the same behaviour (torque spikes) could either bring the fly into the heat or out of it, depending on which quadrant it was in. Instead, the fly had to form two associations. First, it had to associate its behaviour (B) of attempting to fly in the direction of a forbidden quadrant, with the CS (upright T) in that quadrant. Second, it had to associate the CS (upright T) with the aversive property of the US (heat), and learn that the former predicted the latter. This kind of learning is known as operant stimulus conditioning. It proved to be considerably more effective than either classical or pure operant conditioning alone. It was also a more natural way of learning. Whereas in the pure operant conditioning experiments, the fly had to restrict its range and continuously turn to one side to avoid the heat, in flight simulator mode, it simply had to keep its left and right yaw torque spikes within the safe zones.
In switch mode (sw-mode), the fly received two forms of feedback - heat (the US) and a change in the colour (from blue to green) or pattern (from an upright to an inverted T) of the panorama (the CS) - when its yaw torque exceeded the allowable range. For instance, if the fly's yaw torque value fell into the left domain, heat was switched on and the upright T pattern was placed in front; flying in the right domain heat switched the heat off and the arena was quickly rotated by 90 degrees, shifting the inverted T to the front. Thus the fly underwent parallel operant conditioning (a combination of operant and classical conditioning), where it not only had the opportunity to associate its behaviour (B) with a US (heat), but also had the chance to associate a particular display - a colour or pattern (CS) - with the US. Unlike flight simulator mode, the fly had to respond in a very artificial way to avoid the heat: it had to keep flying around in circles to avoid being fried. The requisite behaviour was thus the same as for pure operant conditioning. Brembs wryly comments:
I find it hard to imagine a situation where flying in clockwise circles gets a fly incinerated, but counterclockwise circles prevent that (personal email, 11 August 2003).
In other research (Brembs, 2003), it was shown that flies could stabilise a rotating cylindrical arena by modulating their thrust instead of their yaw torque. Flies learned after a few seconds to adjust their thrust to an arbitrary level (chosen by the experimenter) that corresponded to no rotation of the arena. Less thrust caused the arena to rotate one way (say, counter-clockwise), while more thrust caused it to rotate the other way (clockwise).
Brembs' research findings are summarised in an Appendix.
Brembs (2000) and Brembs and Heisenberg (2001) searched for certain higher-order forms of associative learning that are commonly found in vertebrates undergoing associative learning: blocking, overshadowing, sensory pre-conditioning (SPC) and second-order conditioning (SOC). Research findings for higher-order forms of associative learning are contained in another Appendix. The upshot is that despite similarities between insects and vertebrates, there is no reason to believe that a mind-neutral intentional stance cannot account for the behaviour observed. The following conclusion can be drawn from this research:
L.14 The occurrence of higher-order forms of associative learning in an organism do not, taken by themselves, warrant the conclusion that it has cognitive mental states.
In the next section, however, it will be argued that operant conditioning in Drosophila contains a number of features which, when taken together, do warrant the attribution of agency and mental states (beliefs and desires) to insects.
Back to Chapter 2 Back to: A model of Operant agency *** SUMMARY of conclusions reached References